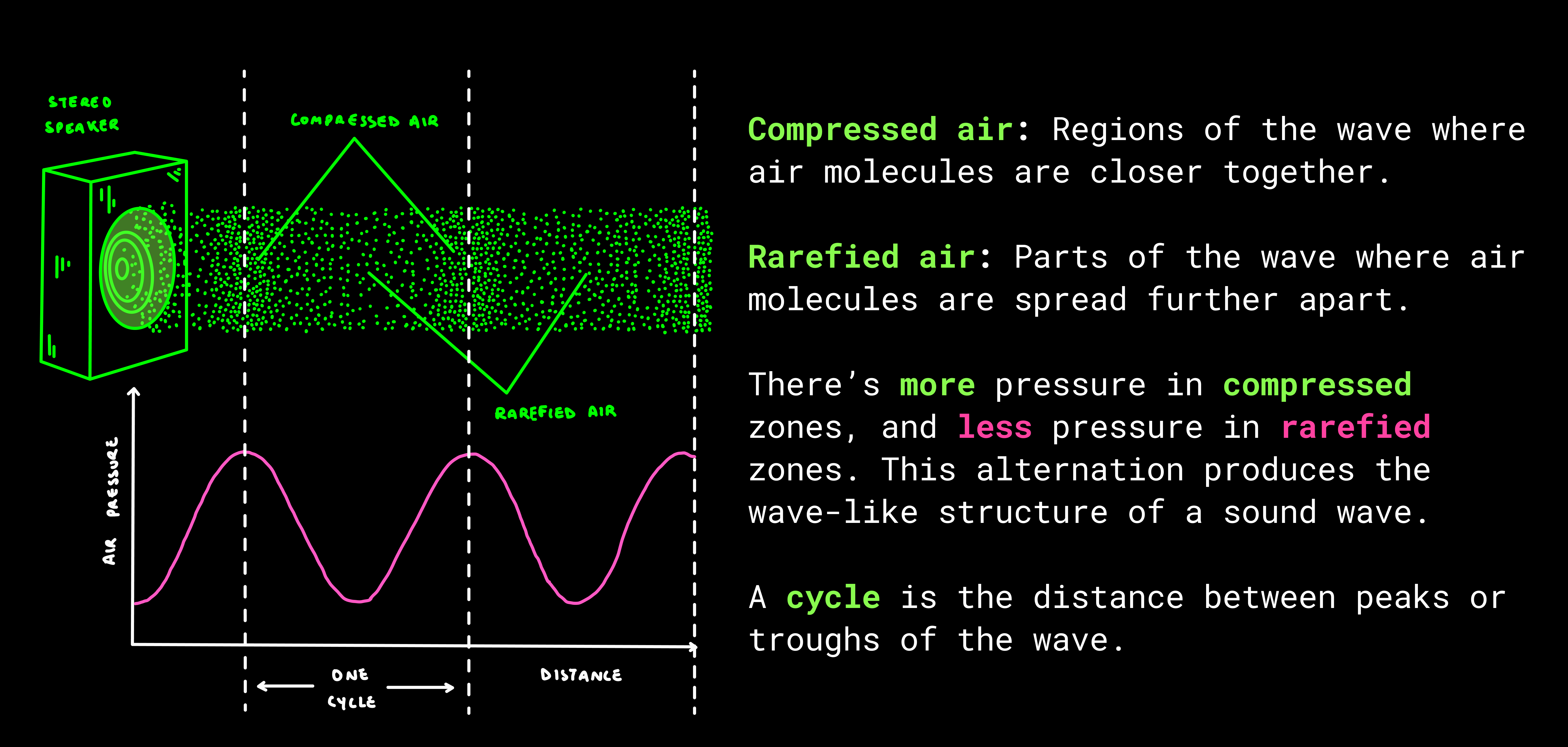
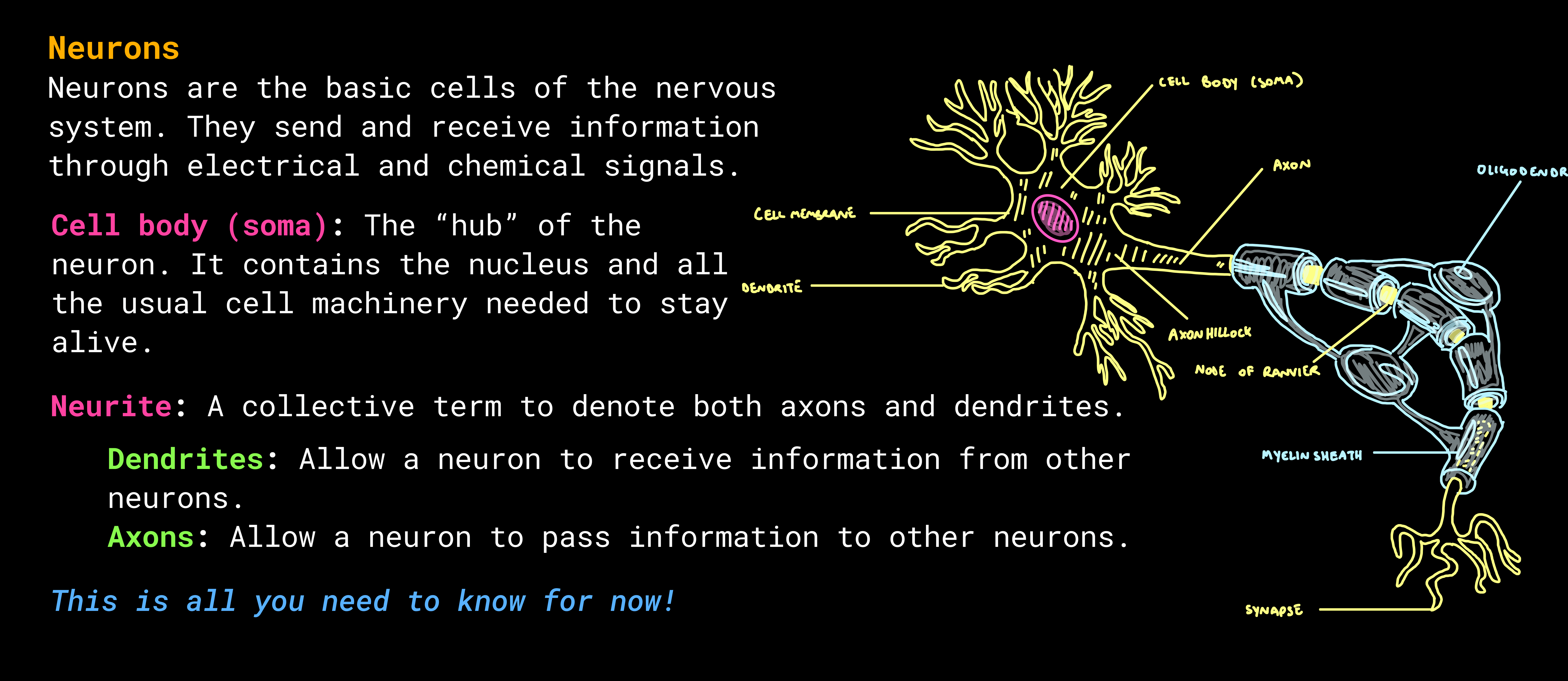
The auditory system works in 3 main stages.
Together, these chambers allow fluid waves to move in carefully controlled patterns.
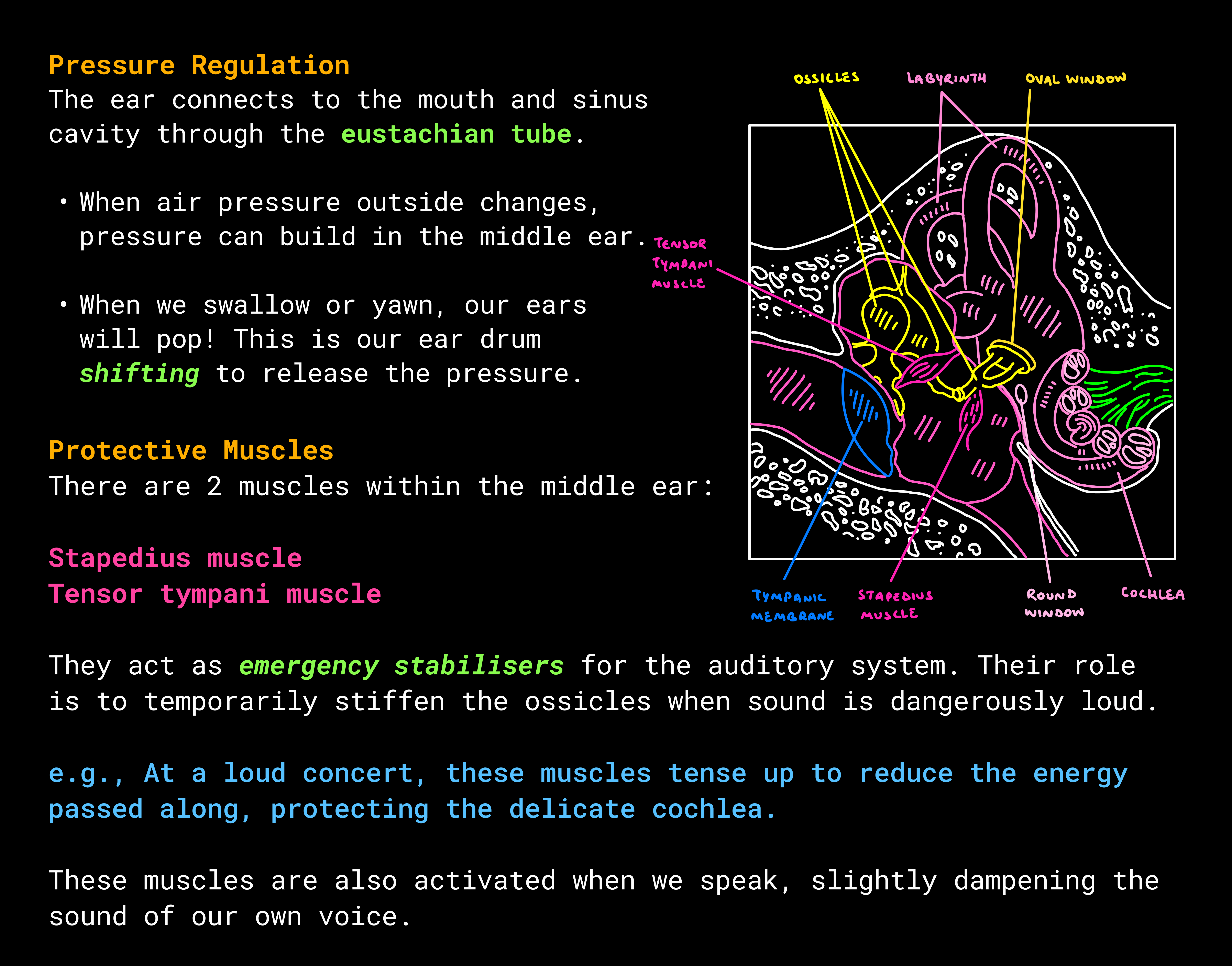
Think of it like a balloon with one thinner patch in its surface. As the balloon fills with fluid and pressure builds inside, that weaker spot flexes outward, releasing strain and equalising the system.
Inside the Cochlea
To understand how the cochlea works, it helps to imagine it uncoiled as a straight tube.
The cochlea is set up to detect sounds at different frequencies.
Along its length runs the basilar membrane, which separates the scala vestibuli and scala timpani.
The basilar membrane isn’t uniform. Each section is tuned to a particular frequency, meaning it vibrates most strongly at that pitch. Mechanical properties determine which frequencies the basilar membrane is sensitive to.
So, the position along the cochlea determines how sensitive that part of the cochlea is to different sound frequencies.
In this way, the cochlea acts like a natural frequency analyser, translating the physical properties of sound into representative neural activity that the brain can interpret.
- High-frequency sounds (i.e birds chirping) create strong vibrations near the base of the cochlea, but fade quickly.
- Low-frequency (i.e. bass drum) travel further along the basilar membrane, peaking near the apex of the cochlea.
This spatial organization is called tonotopy (more on that later)—and it’s what allows us to distinguish between the vast range of pitches we hear every day.
Inside the Organ of Corti
How do vibrations in the cochlea turn into electrical signals?
The answers lies in the Organ of Corti.
The Organ of Corti (as a reminder) is a delicate, specialised structure that sits on top of the basilar membrane.
- As fluid waves move through the cochlea, the basilar membrane vibrates up and down.
- Resting above it is the tectorial membrane, a gelatinous structure that does not move much in response to these waves.
Note: Not to be confused with the tympanic membrane (our eardrum), which vibrates at the very start of the hearing pathway.
- This difference in motion causes hair cells in between to be pushed against the tectorial membrane, deflecting their tiny projections (stereocilia).
Stereocilia are stiff, toothpick-like structures on hair cells that bend in response to vibration.
Hair Cells
Within the Organ of Corti, we have two kinds of sensory receptors—outer and inner hair cells.
Together, they detect vibrations in the basilar membrane and begin the process of turning sound into neural signals.
- Outer hair cells: Their stereocilia are embedded in the tectorial membrane, so they are directly deflected as the basilar membrane moves.
- Inner hair cells: Their stereocilia float freely in the fluid. They respond to the overall fluid motion rather than direct contact.
Note: All hair cells are part of the Organ of Corti. While we often describe them separately as “inner” and “outer” hair cells, they together make up this specialised structure.
Hair cells themselves are not neurons because they don’t fire “all-or-nothing” action potentials.
Instead, they release neurotransmitters in a graded response, proportional to the frequency and amplitude of the incoming sound.
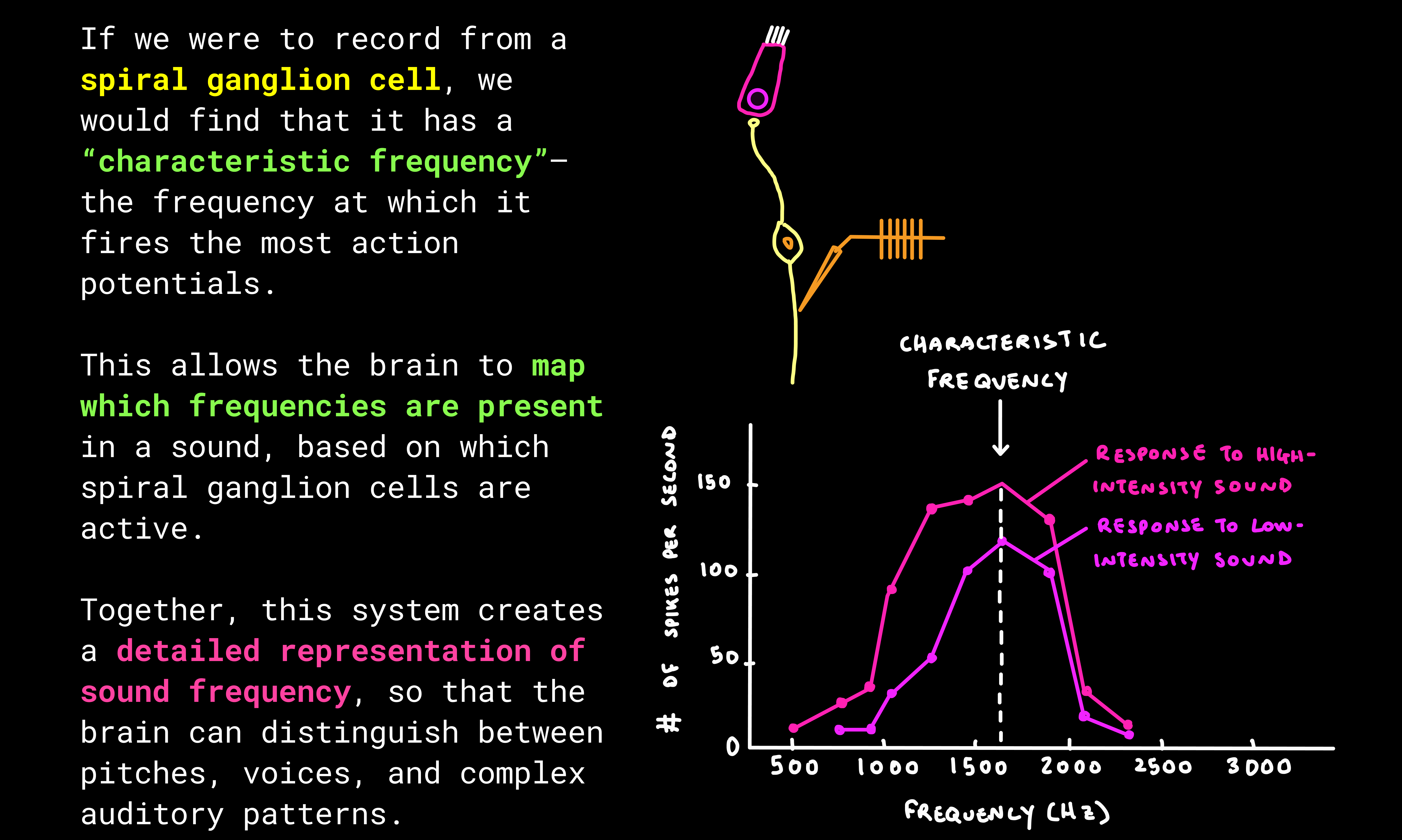
The first true neurons in this pathway are the spiral ganglion cells, which “listen” to the hair cells. Once activated, they fire action potentials and bundle their axons together to form the auditory nerve, carrying sound information to the brain.
Transduction
Below is a step-by-step process on how mechanical vibrations are converted into electrical signals that the brain can interpret.
Resting State
When hair cells are at rest, the basilar membrane is still and the stereocilia are upright. Only a few ion channels are open, so the cell sits near its resting potential.
Step 1: Deflection of Stereocilia
Each stereocilium is connected to its neighbour by tip-link proteins, so when one bends, the entire bundle shifts together.
- As the basilar membrane vibrates, stereocilia bend toward one direction, opening mechanically-gated potassium (K+) channels.
- When they bend the other way, the channels close.
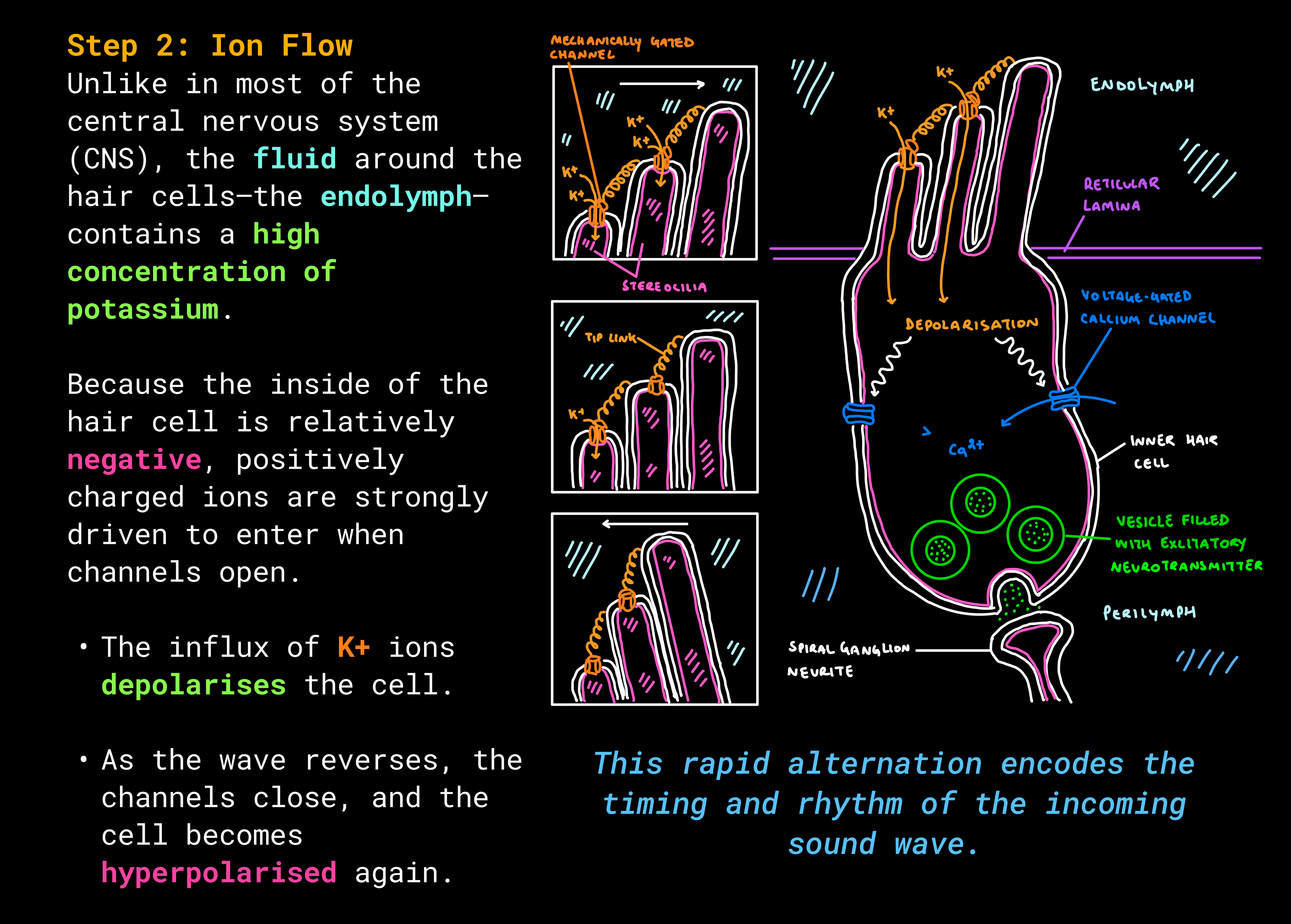
Step 3: Neurotransmitter Release
Depolarisation opens voltage-gated calcium (Ca2+) channels.
- Ca2+ rushes in, triggering vesicles to fuse with the membrane.
- These vesicles release neurotransmitter onto the spiral ganglion cells.
The release is graded—always present at some level, but stronger or weaker depending on the frequency and intensity of the sound.
- Spiral ganglion cells listen to the graded messages of the hair cells and decide whether or not they will fire a binary action potential. They pass this chemical signal into the auditory nerve and onward to the brain.
Recap: Sound wave enters ear → basilar membrane vibrates → stereocilia bend → K⁺ channels open/close → depolarisation / hyperpolarisation of hair cells → Ca²⁺ influx → neurotransmitter release → spiral ganglion fires → auditory nerve → brain.
Differences Between Outer and Inner Hair Cells
Although both types of hair cells detect motion in the cochlea, they connect to spiral ganglion cells in very different ways.
- Many outer hair cells converge onto a single spiral ganglion cell. This means their information is pooled together before being passed on.
- Conversely, we have the opposite arrangement for inner hair cells. One inner hair cell communicates with many spiral ganglion cells. This spreads its information more widely through the pathway.
Why the Difference?
Outer hair cells actually outnumber inner hair cells 3:1.
(Thus, there’s more outer hair cells than inner hair cells)
Yet, most spiral ganglion cells are connected to inner hair cells. Only a small fraction listen to the outer ones.
This is because the two groups encode different aspects of sound.
- Inner hair cells: Specialised for frequency (pitch) detection.
- Outer hair cells: More sensitive to amplitude (volume) intensity.
While amplitude helps us gauge how strong a sound is, frequency is more critical for distinguishing and recognising sounds.
e.g., It’s the change in pitch that lets us tell voices apart or identify a melody.
Hearing Loss
Hearing loss can result from many different causes, depending on where the auditory pathway is disrupted.
Broadly, hearing loss is divided into conductive deafness and sensorineural deafness.
Conductive Deafness
This occurs when sound waves are blocked from reaching the cochlea.
- Common causes: Ear infections, fluid buildup, damaged/calcification of the ossicles, or ruptured ear drum.
In these instances, mechanical vibrations cannot be transmitted effectively through the middle ear.
- Hearing aids are often effective here since they amplify incoming sound waves. Surgery may also help repair ossicles or eardrum damage.
Sensorineural Deafness
This occurs when the cochlea or auditory nerve is damaged.
One of the most common causes is malfunctioning hair cells in the cochlea. In these cases, cochlear implants can help.
- A cochlear implant works by bypassing the damaged hair cells entirely.
It acts like an artificial basilar membrane, directly stimulating the auditory nerve with electrical signals instead of relying on mechanical-to-neural conversion.
These can recover up to ~90% of hearing ability!
However, cochlear implants are not helpful for every type of hearing loss.
- Auditory nerve damage: If the nerve itself is damaged, bypassing the hair cells is not enough, since the pathway to the brain is interrupted.
Mixed Hearing Loss
Sometimes both conductive and sensorineural components are present. For example, someone might have ossicle calcification and cochlear hair cell damage. Procedures may then involve a combination of surgery, hearing aids, or implants, depending on the primary cause.
Cochlear Amplification
Outer hair cells also amplify sound.
On their surface, they contain a special protein called prestin, a voltage-sensitive motor protein.
How It Works
- When the hair cell depolarises, prestin contracts, pushing the cell against the tectorial membrane and boosting the effect of the vibration.
- When the hair cell hyperpolarises, prestin relaxes, allowing the cell to expand and pull away, further enhancing the contrast.

This rapid expansion and contraction acts like a biological amplifier, strengthening the response to sound waves already being detected by the stereocilia.
Why Does This Matter?
Cochlear amplification allows us to detect sounds at very low intensities that would otherwise be too faint to perceive.
It fine-tunes our sensitivity to quiet sounds and subtle differences in pitch.
Clinical note: The drug furosemide can inhibit prestin proteins, removing this amplification mechanism. As a result, the ear becomes less sensitive, particularly to low-intensity sounds.